My overall research centers on the study of an insect order Orthoptera (grasshoppers, crickets, and katydids) and aims at understanding behavioral, ecological, physiological, morphological and molecular evolution in a phylogenetic framework. I believe that there is still a huge gap between process and pattern theories in evolutionary biology and that these two perspectives have to be integrated in order to gain a robust and full understanding of phenotypic evolution. In this sense, I have developed five original and integrative research programs that seek to bridge the gap in various areas of biology.
1. Phylogenetic systematics of Orthoptera (Insecta)
Orthoptera is a fascinating group of insects that contains grasshoppers, locusts, katydids, and crickets, well known for their ability to jump and acoustic communication. Despite its familiarity, the phylogenetic relationships within Orthoptera are poorly understood and the classification is not stable because of a long history of conflicting taxonomic schemes based on different morphological traits.
My research aims at improving the current status of the systematics of Orthoptera and it covers all areas of systematic biology including alpha-level taxonomy [Song 2006] and revision [Song 2004b], phylogenetic analyses at various levels using both traditional data such as external and internal morphology [Song 2004c], behavioral and ecological characters [Song and Wenzel 2008a] as well as modern molecular data including the whole mitochondrial genome. In 2008, I received a research grant from the U.S. National Science Foundation (DEB-0816962) as a Principal Investigator to reconstruct a comprehensive phylogenetic hypothesis of Orthoptera, based on complete mitochondrial genomes and five nuclear genes (18S, 28S, wingless, CAD, arginine methyltransferase).
So far, two major collecting trips (South Africa and Australia) and collaborations with the orthopterists worldwide have provided DNA-grade tissue samples to have the most complete taxon sampling of about 400 species which covers every known family and most subfamilies. I have involved several undergraduate students in sequence data generation for them to gain research experiences and this collaboration has already generated about one-third of data needed for this project. Through this project, I will propose testable phylogenetic hypotheses for the major lineages of Orthoptera, which will serve as a foundation for further systematic research for this interesting group of insects. A number of international collaborative projects have been initiated from this project with the researchers from France, Germany, Argentina, Australia, and China. My long-term goal is to train the next generation of orthopterists through an active systematics program because Orthoptera, unlike other major taxonomic groups, lacks taxonomic expertise and there are only a handful of active specialists around the world.
2. Evolution of nuclear mitochondrial pseudogenes
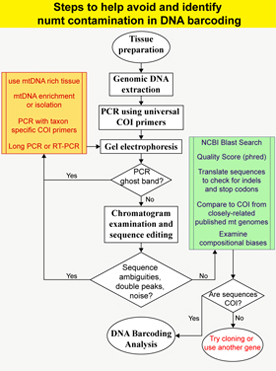
Nuclear mitochondrial pseudogenes (numts) are non-functional copies of mtDNA in the nuclear genome which have been found in the major clades of eukaryotic organisms. Among insects, Orthoptera, especially grasshoppers, are known to have exceptionally high numbers of numts although the exact mechanism for nuclear integration is not well understood. The presence of numts poses a challenge for PCR-based mitochondrial systematics because inadvertent coamplification or preferential amplification of numts can occur, thereby compromising the orthology assumption of characters. This line of research stemmed from my own struggle with generating mitochondrial sequences from grasshoppers, which has serendipitously transformed into an exciting area of research. My first approach was to document the effect of numt coamplification in mitochondrial systematics and as a case study, I examined the DNA barcoding initiatives, which relies on a portion of
mitochondrial cytochrome c oxidase I (COI) gene to correctly identify species [Song et al. 2008]. I showed that standard barcoding primers can easily coamplify numts of COI gene from diverse lineages within arthropods (insects and crustaceans), that some numts are difficult to identify because they do not have characteristic stop codons or indels, and that numt coamplification can potentially overestimate the number of species in DNA barcoding. Currently, as a part of the NSF project, I am documenting a similar phenomenon in other genes within mtDNA from 25 major lineages of Orthoptera. One of the most interesting findings to date is the idea that numts can actually be used as phylogenetically informative characters because they represent ancient forms of mtDNA or molecular fossils, silently lodged in the nuclear genome. I have recently tested this idea using numt sequences from 21 species of a grasshopper genus Schistocerca and the resulting manuscript has been submitted. The research on numts has a number of potentials that can lead to exciting discoveries, and I intend to continue and expand this research program.
3. Mitochondrial genomics
Now is the age of very large molecular data in systematics. However, our ability to analyze large and complex molecular data is severely outpaced by our ability to generate such data. My research aims at bridging this gap in the context of mitochondrial genomics. Mitochondrial genomes (mtgenomes) are the smallest extant organellar genome, which in insects encode for 13 protein-coding, 22 tRNA and 2 rRNA genes with an average size about 15,000 bp. It is now technically feasible to sequence the mtgenome of a given organism in its entirety within a short period of time. The complexity of the genome structure and the large yet manageable size of the genome make mitochondrial genomics an ideal model system for exploring various challenges of today’s molecular systematics. As a postdoctoral research fellow of an NSF-funded AToL Beetle Tree of Life Project (PI: M.F. Whiting), I generated complete mtgenome sequences for 72 key beetle species and I was involved in developing a bioinformatics tool that can aid genome annotation and data management [MOSAS]. Currently, I am generating complete mtgenome sequences for 25 major taxa in Orthoptera as a part of my ongoing NSF project. The availability of mtgenomes from diverse lineages within a given taxonomic group allowed me to study genome evolution in a comparative context. In Coleoptera, I examined the evolution of genome structures, atypical stop codons, and transfer RNAs [Sheffield et al. 2008] and in Orthoptera, I studied lineage-specific gene rearrangements in a phylogenetic framework [Fenn et al. 2008]. My research also focused on how best to analyze mtgenome data as a phylogenetic marker for deep-level relationships. So far, my research shows that mtgenome data are often highly affected by the past molecular events, resulting in patterns such as among-site rate variation and base compositional heterogeneity and that incorrect phylogenetic inference is inevitable when such systematic bias is not accounted for [Sheffield et al. 2009]. I am currently exploring various ways to overcome systematic bias in phylogenetic reconstruction. The future direction of this research will be focused on investigating how mtgenome data behave in phylogenetic reconstruction which is yet to be fully understood.
4. Evolution of insect male genitalia
Among animals with internal fertilization, many species have species-specific male genitalia with morphological divergence among closely related species that is often dramatic and complex. This pattern is especially evident in insects, and male genitalia are considered one of the most important and useful species-diagnostic characters in insect systematics. Recent theoretical developments in genital evolution show that male genitalia are under sexual selection and evolve rapidly, and in some cases, sexual selection on male genitalia is thought to be a driving force behind speciation and diversification. The study of genital evolution is one of the most active areas of research in evolutionary biology today. I became naturally interested in insect male genitalia early on in my career while I was conducting a taxonomic study of grasshoppers, which relied heavily on the morphology of male phallic structures [Song 2004a]. That interest has eventually transformed into an active research program, which is highly integrative in a sense that it covers several different areas
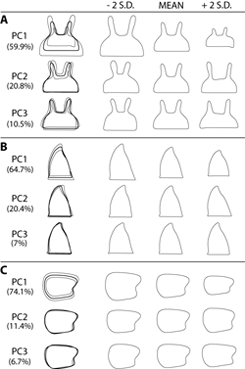
of biology including, developmental biology [Song 2004a], functional and comparative morphology [Song and Wenzel 2008b], [Song 2009], ecology and systematics [Song and Bucheli 2009]. Recently, I became very interested in studying the role of sexual selection in shaping genital diversity, and therefore species diversity. For instance, if a lineage with high genital diversity amongst its members has high rates of diversification, inferred from a phylogeny, sexual selection may explain the genital evolution in that lineage. Many lineages within grasshoppers exhibit acoustic or visual pre-mating courtship behaviors, which are presumably also under sexual selection, but it is not clear whether male genitalia are still under sexual selection in these lineages. In order to test whether sexual selection is a major factor in the diversification of grasshoppers, it is important to quantify the amount of shape variation in genital morphology from many lineages with different mating strategies in a robust phylogenetic framework. For the past two years, I visited four major museums rich in Orthoptera (ANSP, MNHN, BMHN, and ANIC) and borrowed over 1000 specimens representing all major lineages of grasshoppers. Over the next three years, I intend to dissect and quantify male genital morphology using high-resolution imaging and geometric morphometrics and to test the hypothesis in light of the molecular phylogeny which is currently being produced.
5. Evolution of phenotypic plasticity in swarming locusts
Locusts are grasshoppers that can form dense migrating swarms through a phenomenon known as locust phase polyphenism [see this website for more detail]. There had been a number of advances made in the areas of behavioral ecology, physiology, and biochemistry which helped understand the mechanism of locust swarming, but the evolutionary relationship between locusts and non-swarming relatives was not well-understood, preventing scientists to see the big picture. The results from my dissertation based on a morphological phylogenetic analysis
suggest that the locust species within a given lineage do not necessarily form a monophyletic group, indicating that locust phase polyphenism must have evolved multiple times [Song 2004c, Song 2005, Song and Wenzel 2008a]. Moreover, locust species are often closely related to sedentary species that are capable of expressing density-dependent phenotypic plasticity, which has resulted in a novel hypothesis that the locust swarming is a result of a stepwise assembly of phylogenetically conserved phenotypic plasticity, evolved throughout the diversification of grasshoppers. I plan to expand this line of research by developing a full-scale comparative study to empirically investigate the expression of phenotypic plasticity in the phylogenetically close species which can ultimately pinpoint what makes grasshoppers become locusts from both pattern and process perspectives.